功能食品蛋白是一类具有改善食品质构、增强风味、赋予色泽或调节生理活性等特殊功能的蛋白质。根据其核心功能差异,可分为酶制剂、呈味蛋白、色素蛋白和乳蛋白等。随着居民生活水平提升与消费结构升级,市场对食品营养价值和感官品质的需求持续增长。据Grand View Research统计,2023年,功能食品蛋白市场规模已达65亿美元(约465.9亿元),并预计以5.3%的年复合增长率持续扩张[1]。然而,除食品酶外,呈味蛋白、色素蛋白和乳蛋白等多种功能食品蛋白的生产方式仍高度依赖畜牧养殖或植物提取,存在生产效率低、占地面积大、温室气体排放多等问题。据联合国粮农组织统计,全球约有18%的温室气体排放源于畜牧业。同时,食品酶的合成也面临生产成本高和产物活性低的问题。因此,急需变革功能食品蛋白的传统制造模式。
近年来,合成生物学等前沿技术的突破性进展,为食品产业的转型升级提供了核心技术支撑[2]。作为融合生物信息学、工程学与分子生物学的交叉学科,合成生物学通过对生物系统的理性设计与定向改造,能够显著提升目标产物的合成效率。利用合成生物学技术构建微生物细胞工厂,实现功能食品蛋白的绿色高效生物制造,是践行“大食物观”和国家“双碳”目标的重要途径[3]。例如微生物法的蛋白合成效率是种植和养殖业的102~103倍,而占地面积仅需种植和养殖业的1/100~1/10,温室气体排放量为种植和养殖业的20%~50%[4]。此外,合成生物学技术还赋能酶分子的精准设计改造,通过重构蛋白质的三维结构,显著提高食品酶的催化活性。鉴于此,本文以食品酶、呈味蛋白、色素蛋白及乳蛋白为例,总结了它们在食品领域的功能价值与应用,系统分析了功能食品蛋白生物制造的发展现状,并展望了其未来所面临的机遇与挑战。
1 食品酶研究领域
食品酶是一类能够加速食品加工过程并提升产品品质的生物催化剂。据Introspective Market Research统计,2024年全球食品酶市场产值达25.5亿美元(约合人民币182.9亿元),预计到2032年增长至41.6亿美元(约合人民币298.4亿元)[5]。如何高效获取催化性能优异、稳定性突出的优质酶是食品绿色生物制造所面临的核心挑战之一。近年来,合成生物学与酶工程技术的快速发展为食品酶的高效制造提供了关键技术支持。本节以转谷氨酰胺酶 (肉类或乳制品加工的关键酶)和醇脱氢酶、醛脱氢酶 (风味与保鲜功能酶)为例,阐述食品酶生物制造的研究进展与应用现状。
1.1 转谷氨酰胺酶
转谷氨酰胺酶(transglutaminase,TGase)属于酰基转移酶家族,广泛存在于动物、植物和微生物中。其核心功能是催化蛋白质分子间或分子内形成ε-(γ-谷氨酰基)赖氨酸异肽键,介导蛋白质交联反应[6-7]。在食品加工中,TGase是重塑食品质构的“分子缝合器”,能有效调控蛋白质基产品的溶解度[8]、持水能力(water holding capacity,WHC)[9]、黏度[10]、弹性[11]及凝胶性[12],因而在肉制品、乳制品和植物蛋白制品等领域应用广泛[13]。
TGase的生产途径主要包括两类:生物提取法和微生物发酵法[14]。早期,TGase的生产主要依赖动物器官提取 (如豚鼠肝脏、牛血浆、鱼类卵巢),需经组织破碎、离心、盐析、层析等多步纯化[15],存在原料稀缺、成本高昂等问题[16-17]。在植物体中,TGase主要存在于种子 (如绿豆、向日葵)、块茎 (如马铃薯)或叶片 (如菠菜)中,但其含量极低,且提取过程易受植物次生代谢物如单宁、多酚等抑制干扰,商业价值较低[6]。
1989年,Ando团队[18]首次鉴定了茂源链霉菌(Streptomyces mobaraensis)来源的TGase。此后,研究者陆续在吸水链霉菌(S. hygroscopicus)[19]、谷氨酸棒杆菌(Corynebacterium glutamicum)[6]、大肠杆菌(Escherichia coli)[20]中发现了天然的TGase。相较生物提取法生产TGase,微生物发酵法生产TGase具有合成效率高、成本低等优点。然而,天然微生物宿主普遍存在TGase本底表达量低的问题。因此,需对天然宿主进行遗传改造以提高TGase合成效率。在微生物中,TGase常以无活性的酶原形式 (pro-TGase)分泌,经激活蛋白酶活化后方可转化为活性酶[21]。Yin等[22]发现在培养基中添加NH4+能显著提升S. mobaraensis中激活蛋白酶的活性。通过系统优化NH4+的添加剂量与时间,pro-TGase激活效率大幅提高,最终使TGase的产率提升2.1倍,生产周期缩短43%。Yuan等[23]通过增加S. mobaraensis中TGase编码基因的拷贝数、使用高强度的启动子驱动TGase的表达和优化TGase的信号肽等改造策略,构建了一株TGase高产菌株,在摇瓶和1 000 L发酵罐中的酶活分别达56.43 U/mL和63.18 U/mL,是目前已报道的最高酶活。
此外,由于S. mobaraensis等TGase天然合成菌株遗传改造困难,因此有研究尝试在酿酒酵母(S. cerevisiae)等模式工业菌株中异源表达TGase,以期获得TGase的高表达菌株。S. cerevisiae作为食品级安全模式真核微生物,能够天然合成pro-TGase的激活剂谷胱甘肽,有利于活性TGase的表达。Hirono-Hara等[24]前期基于代谢工程策略成功构建了一株能够高产谷胱甘肽的酿酒酵母工程菌株;基于该底盘细胞,研究者成功建立以脱脂乳清为碳源的TGase生产方式[25] (图1)。
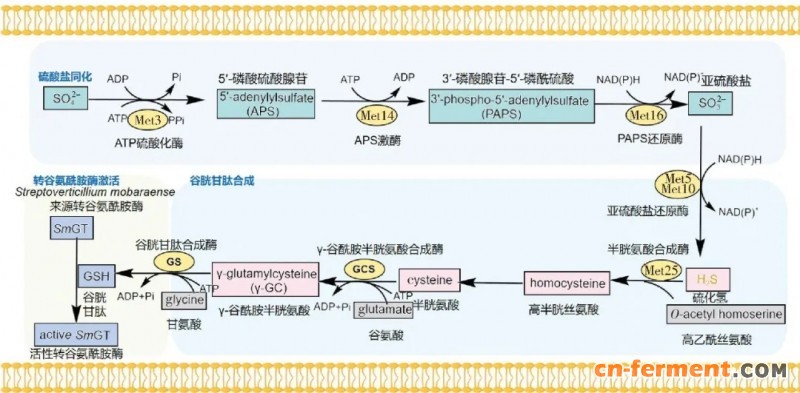
图1 以酿酒酵母为宿主合成活性TGase的代谢过程
除表达量较低外,天然TGase还存在热稳定性差和催化效率低等问题。为解决上述问题,Yang等[26]以S. mobaraensis来源的TGase为研究对象,结合随机突变、分子对接和分析动力学模拟等酶工程技术,成果获得了一个催化活性和热稳定显著提高的TGase突变体FRAPD-TGm2A,其在60 ℃条件下的半衰期达132.38 min,较野生型提升84%,比酶活达79.15 U/mg,较野生型提升21%。
1.2 醇脱氢酶和醛脱氢酶
醇类和醛类化合物是环境、日化、食品加工及畜禽养殖等领域异味污染的核心组分。作为生物催化体系的关键成员,醇脱氢酶 (alcohol dehydrogenase, ADH)与醛脱氢酶 (aldehyde dehydrogenase, ALDH)凭借其高效性、高特异性、温和的反应条件及环境友好性,在异味消除技术中展现出显著优势[27](图2)。 ADH主要催化伯醇或仲醇可逆氧化为醛或酮,对短链至中链脂肪醇 (如具腐败气味的戊醇)及芳香醇 (如苯甲醇)具有广泛活性[28];而ALDH则可将醛类氧化为羧酸,对清除低感官阈值、高挥发性的醛类异味尤为有效[29]。因此,ADH/ALDH体系被广泛应用于食品工业,如大豆除腥[30]、乳制品脱膻、肉制品保鲜[31]及酒类除杂醇[32]等,并延伸至食品加工环境异味治理领域,例如肉制品厂废水处理与烘焙车间空气净化等。
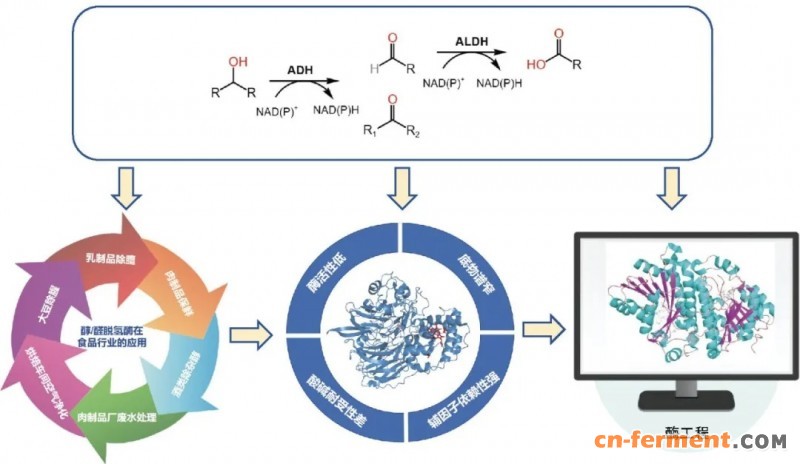
图2 醇/醛脱氢酶在食品行业中的应用现状
ADH/ALDH的生物制造技术经历了从天然提取到智能设计的跨越式发展。早期主要依赖天然菌株筛选,并通过发酵优化提升酶产量,但存在酶活低、杂质多及底物谱窄等局限。随着基因工程技术的突破,研究者得以从极端微生物中克隆ADH/ALDH基因,并在E.coli、S.cerevisiae等工程宿主中实现高效表达,显著提升了酶的产量与纯度。进入21世纪后,蛋白质工程成为核心技术驱动力,通过理性设计、定向进化、半理性设计以及辅因子工程等策略,显著优化了酶的性能。例如,江森等[32]以S. cerevisiae为出发菌株,构建了过表达乙醛脱氢酶ALD6的重组菌株α-ALD6。在传统黄酒发酵工艺条件下,可以促进乙酸、降低高级醇的生成,其中乙酸产量是出发菌AY12α的2.92倍,总高级醇产量下降72.04%。
天然来源ADH/ALDH存在显著局限性,如酶活低、底物谱狭窄、酸碱耐受性差等。针对酶活性低的问题,Zheng等[33]对E.coli来源的醇脱氢酶AdhP进行了改造。他们首先基于计算方法鉴定出12个关键残基,并构建了单点突变库,筛选获得24个对香草醛活性提升的阳性突变体。随后,对有益突变位点进行组合突变,获得了一个催化效率提升8.5倍的6点突变体M6。Xu等[34]开发了1套整合虚拟筛选、理性设计与双评分机制的高效乙醛脱氢酶 (alcohol dehydrogenase, ADA)工程化改造策略。首先构建了包含5 000个Dickeya parazeae ADA (DpADA)同源酶的虚拟库,利用深度学习预测酶的Kcat值,并结合分子对接筛选,最终从Buttiauxella sp. S04-F03中鉴定出高活性候选酶ADA6。该酶催化乙醛至乙酰辅酶A的转化率达到57.6%,活性较基准酶提升了14.1倍。为进一步提升该酶热稳定性,研究者通过折叠工程 (FoldX能量计算指导)获得了突变体P443C,其经热处理后的残余活性显著提高了80.7%。基于分子动力学模拟揭示的底物通道关键瓶颈位点I440,研究团队建立了融合结构调控 (空间位阻)与电子效应优化的双评分系统,并据此设计出双突变体P443C/I440T,最终实现了93.2%的转化效率。
2 呈味蛋白研究领域
呈味蛋白是一类能够增强、修饰或掩盖食品风味的蛋白质。它们通过作用于舌头上的味觉受体,直接引发或调控味觉感知。基于此特性,呈味蛋白能够在维持食品原有风味品质的同时,显著降低加工过程中糖、盐等配料的用量,从而满足现代消费者对“减糖不减甜”、“减盐不减咸”等健康食品的核心诉求,展现出广阔的应用前景。
2.1 甜味蛋白
甜味蛋白是一类源自天然植物的大分子蛋白质类甜味剂,甜度约为蔗糖的100~3 000倍。随着生活水平的逐步提高,蔗糖等传统糖类甜味剂过量摄入所引发的糖尿病、肥胖症及龋齿等健康问题日益突出;与此同时,阿斯巴甜、三氯蔗糖等化学合成甜味剂的安全性也备受争议。相比之下,甜味蛋白凭借其天然来源和富含氨基酸等营养特性,成为兼具安全性与功能性的理想甜味剂选择,在新型甜味剂开发领域展现出巨大的应用潜力,因此备受关注。
目前已鉴定的主要甜味蛋白有8种:莫内林(monellin)、索马甜蛋白(thaumatin)、巴西甜蛋白(brazzein)、仙茅甜蛋白Ⅱ(neoculin)、倍他丁(pentadin)、非洲奇果蛋白(miraculin)、马槟榔甜蛋白(mabinlin)和仙茅甜蛋白Ⅰ(curculin)[35-41]。根据其味觉特性可划分为3类:1)莫内林、索马甜、巴西甜蛋白、倍他丁和马槟榔甜蛋白本身具有甜味;2)非洲奇果蛋白与仙茅甜蛋白Ⅰ虽自身无甜味,但具有甜味调节功能 (可将酸味转化为甜味,通常也归类为甜味蛋白);3)仙茅甜蛋白Ⅱ则兼具上述两类特性。下面重点阐述目前微生物合成研究最为深入的3种甜味蛋白的最新进展 (图3)。
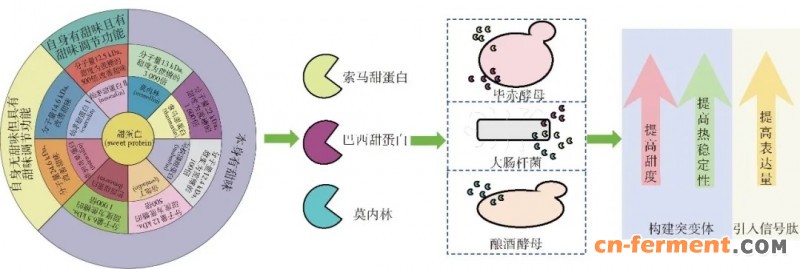
图3 3种甜蛋白生物合成的细胞工厂构建及其改造策略
莫内林是一种来源于非洲热带植物应乐果(Dioscoreophyllum cumminsii Diels)果实的甜蛋白,甜度约为蔗糖的3 000倍[42]。Chen等[43]利用基于质粒pET-22b构建了莫内林表达载体,并导入E. coli,发酵后莫内林产量达43 mg/g(以细胞干质量计)。Aghera等[44]同样以E. coli作为宿主,但使用质粒pET-DUET表达莫内林编码基因,最终产量达 40 mg/L。 Chen等[45]以S. cerevisiae为表达宿主,通过优化启动子和信号肽,使莫内林产量达到 410 mg/L。 Liu等[46]通过在S. cerevisiae基因组中多拷贝整合莫内林编码基因,获得一株产量达675 mg/L的莫内林生产菌株。为了增强重组莫内林的甜味强度和热稳定性,研究者开发了多种突变体。其中,突变体Q28K/C41S/Y65R和E23Q/Q28K/C41S/Y65R的甜度分别提高1.5倍和2.5倍[47],且突变体E23Q/Q28K/C41S/Y65R的热稳定性显著提升,熔解温度达到77.8 ℃。
巴西甜蛋白(亦称植物甜蛋白)来源于西非植物忘忧果(Pentadiplandra brazzeana Baillon)果实的植物甜蛋白,是目前已知的分子质量最小(约6.5 kDa)且热稳定性最高的天然甜味蛋白。Berlec等[48]基于E. coli的密码子偏好性,优化了植物甜蛋白的编码基因,并将其导入E. coli进行异源表达,植物甜蛋白产量达到35 mg/L。进一步对纯化后的重组蛋白进行感官评估,发现其甜度约为蔗糖的1 000倍[48]。赵红玲等[49]将植物甜蛋白编码基因导入毕赤酵母(Pichia pastoris) GS115,菌株发酵后甜蛋白产量达 385 mg/L。为进一步提高其甜味性能,韩国学者构建了H31R/E36D、H31R/E41A和E36D/E41A三种双突变体,其甜度均显著高于野生型植物甜蛋白,其中前两者的甜味强度超过野生型6倍。在此基础上构建的三重突变体H31R/E36D/E41A甜度进一步提升至野生型的12倍[50]。
索马甜蛋白源自西非热带植物西非竹芋(Thaumatococcus daniellii Benth)果实,分子量约为22 kDa,甜度约为蔗糖的1 600倍。Masuda等[51]以pPIC9K为载体在P. pastoris中表达索马甜蛋白,产量约为25 mg/L。由于索马甜蛋白已实现产业化,且表达体系较为复杂,目前关于其分子工程改造的研究相对较少。Masuda发现21位碱基的突变(D21N)可以显著提升索马甜蛋白的甜度;此外,多个带正电荷的表面氨基酸位点在突变后,蛋白甜度显著下降,提示这些残基可能在蛋白与甜味受体的识别和结合过程中发挥关键作用[52]。
2.2 鲜味肽
鲜味肽是一类具有显著鲜味的小分子肽,分子量在150~3 000 Da,广泛存在于多种食品基质中。其制备方式主要包括天然来源提取与化学合成两类路径。早在1978年,Yamasaki首次从牛肉肉汁中鉴定出一条具有鲜味的八肽,氨基酸序列为Lys-Gly-Asp-Glu-Glu-Ser-Leu-Ala[53]。该八肽后被命名为牛肉风味肽(BMP),分子质量为847.39 Da,等电点为4.1[54-56]。当前,BMP的获取方法主要包括化学合成法、酶解法和微生物发酵法。其中,化学合成法产率有限且存在试剂残留风险;酶解法成本高、纯化复杂;相较而言,微生物发酵法合成BMP具有产量高、成本低和易纯化的优势[57]。
Lerch等[58]将BMP编码基因克隆至含信号识别肽序列的表达载体pNS2,分别在E. coli及酵母宿主系统中实现了BMP胞外分泌表达。天津科技大学王艳萍团队对BMP表达菌株构建及其风味生成机制进行了系统研究。其工作内容包括BMP编码基因在P. pastoris[59]和S. cerevisiae[60]中的异源表达、表达系统的优化、生物反应器发酵条件优化,以及发酵产物经美拉德反应生成牛肉风味香精的工艺开发[61-63]。此外,张崟等[64-65]针对重组与天然BMP之间的结构与风味差异,首先构建了E. coli表达系统以实现重组BMP的表达,并结合纯化分析和感官评价手段,深入探讨了其结构基础与呈味效应之间的关系,为后续风味肽的理性设计提供了理论支持。
3 色素蛋白研究领域
色素蛋白是一类具有特征性色彩的功能性蛋白质,因其天然、安全、无毒且色泽鲜艳,在食品工业中具有重要应用价值。相比传统的人工合成色素不仅具备优异的着色性能,通常还兼具抗氧化等生理活性。因此,色素蛋白有望成为天然色素的理想替代品,不仅契合现代消费者对“清洁标签”和健康营养的诉求,也为功能性食品及高端生物制品的开发提供了优质原料,应用前景广阔。
3.1 血红蛋白
血红蛋白(hemoglobin,Hb)是一类以铁卟啉为辅基的高分子蛋白质,其核心结构由亚铁离子与原卟啉IX结合形成的血红素和两对不同的珠蛋白链 (α链和β链)组成,广泛存在于原核与真核生物细胞中,参与氧气运输与储存、一氧化氮双氧合及细胞内pH调节等多种生理过程[66]。随着现代生物技术的发展,血红蛋白的应用已超越其传统生理功能,拓展为具有工程化潜力的多功能蛋白,广泛应用于医药和食品领域。例如,在植物性肉类替代品中添加血红素蛋白可模拟肉类的风味与色泽[67]。
基于氧化锌-介孔碳聚合材料的固相萃取技术虽可用于从血液中分离血红蛋白[68],但该方法存在原料有限、成本高昂、产量低及规模化困难等问题。相比之下,微生物发酵法合成血红蛋白在产量和成本等方面更具优势[69]。E. coli因其操作简便、培养成本低,被广泛应用于重组血红蛋白生产,但需克服包涵体形成、α亚基不稳定以及血红素供应不足等问题。因此,研究人员通过密码子优化[70]、融合蛋白标签[71]增强珠蛋白亚基的可溶性表达,采用短肽(Gly)串联α亚基并共表达α-血红蛋白稳定蛋白[72-73],有效克服α亚基构象不稳定难题,针对大肠杆菌内源血红素合成途径受限问题,通过外源添加血红素、过表达血红素转运蛋白[74],成功实现人血红蛋白的高效体内自组装,表达量可达6.4 g/L[75](图4)。
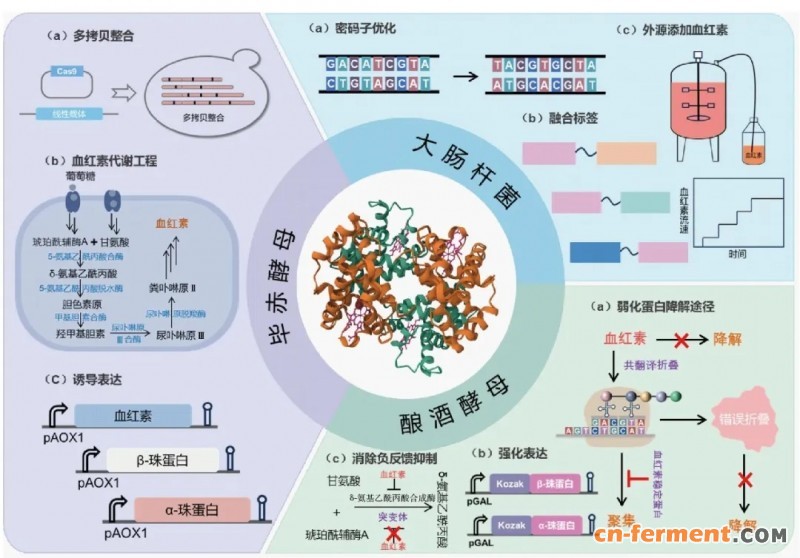
图4 基于合成生物学的血红蛋白细胞工厂调控策略
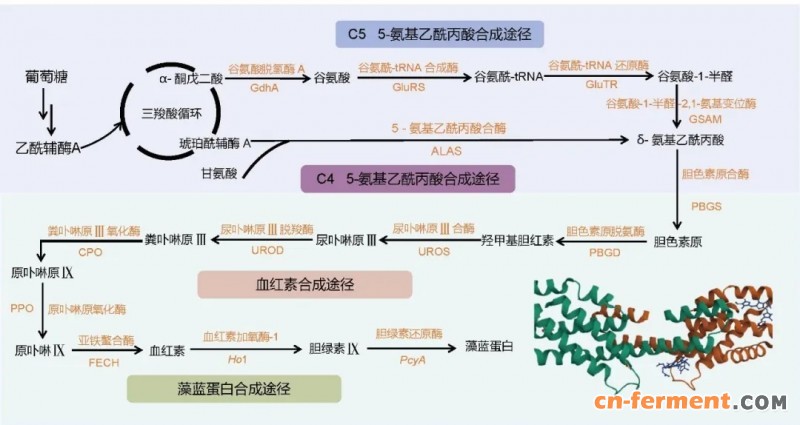
图5 藻蓝蛋白生物合成途径
然而,大肠杆菌在食品应用中存在安全隐患,且当前研究多集中于动物源血红蛋白,需经适当折叠才具活性。相比于大肠杆菌,酵母等真核表达系统更适合用于合成血红蛋白。前期,研究人员主要利用S. cerevisiae合成人血红蛋白,通过改造S. cerevisiae异源蛋白降解路径、过表达人-Hb α亚基稳定蛋白AHSP,血红蛋白积累达总蛋白的18%[76],但此产量难以满足实际应用需求。随后,通过调控转录水平与翻译效率实现珠蛋白亚基的均衡表达,同时解除血红素对 Hem1p(δ-氨基乙酰丙酸合成酶)的负反馈抑制效应,并增强血红素合成通路中关键酶的表达量,系统性提升珠蛋白和血红素协同合成效率,合成了高活性血红蛋白,包括人血红蛋白(174.2 mg/L)、猪血红蛋白(203.9 mg/L)和牛血红蛋白(258.2 mg/L)[77]。尽管如此,S. cerevisiae较低的蛋白分泌能力仍制约其工业化应用。
P. pastoris因其高效的蛋白表达与分泌能力,成为血红蛋白合成领域更具前景的底盘细胞。Shao等[78]基于P. pastoris系统,通过多拷贝整合LegH基因与代谢工程改造血红素合成途径,成功开发出高效分泌大豆血红蛋白的工程菌株。经发酵条件优化,在10 L规模发酵罐中目标蛋白产量提升至3.5 g/L。 Impossible Foods公司则利用甲醇诱导型AOX1启动子,强化血红素合成通路及大豆血红蛋白编码基因表达,并调控AOX1转录水平,同时优化放大培养工艺参数与代谢网络适配性,成功构建了血红蛋白的微生物制造平台[79]。
3.2 藻蓝蛋白
藻蓝蛋白(phycocyanin,PCB)是一类由脱辅基蛋白亚基和藻蓝胆素(phycocyanobilin,PCB)发色团通过硫醚键共价结合而成的捕光色素蛋白,天然存在于蓝藻和藻类的光系统中[80]。其基本结构为α/β异源二聚体,每个亚基通过其保守半胱氨酸残基特异性结合1个四吡咯环结构的藻蓝胆素分子[81]。凭借其亮蓝色泽、抗氧化及抗炎特性,PCB在多个领域具有广泛应用前景。2014年,美国食品药品监督管理局批准PCB作为天然食品着色剂,可用于糖果和烘焙糕点等食品[82-83]。
目前,PCB主要从螺旋藻中通过反复冻融结合超声波破碎细胞,再经硫酸铵盐析进行分离纯化获取[84]。然而,该方法存在超声处理量小、纯化步骤烦琐耗时等局限,难以满足工业化生产需求。鉴于此,利用微生物合成PCB的策略日益受到关注 (图 5)。 早期研究利用pET28a质粒在E. coli中异源表达Ho1和PcyA基因,并以外源血红素作为底物合成藻蓝蛋白。但受限于ho1和pcyA基因表达量低及血红素的反馈抑制,PCB产量仅为3 mg/L[85]。随后,通过采用双顺反子启动子表达ho1和pcyA基因并增加其拷贝数,PCB产量提升至6.64 mg/L[86],此结果证实ho1和pcyA催化的两步反应是PCB合成的限速步骤。为提升产量,研究者利用携带诱导型T7lac启动子的高拷贝数质粒,以2∶1的比例共表达ho1和pcyA基因,并直接添加前体底物5-氨基乙酰丙酸 (ALA),将PCB产量提高至28.32 mg/L[87]。为突破产量瓶颈,研究人员采用酶分子改造与代谢调控协同策略提升PCB的合成效率。首先筛选不同来源的Ho1和PcyA同源酶,经高通量定向进化获得高效突变体,利用DNA支架技术降低酶分子间空间位阻,并适度强化血红素合成关键基因hemB和hemH表达。经5 L发酵罐工艺优化,PCB产量提升至184.20 mg/L[88]。
4 乳蛋白研究领域
乳蛋白是存在于动物乳汁中的优质蛋白质,主要包括酪蛋白和乳清蛋白 (乳铁蛋白、骨桥蛋白、乳白蛋白等)两大类,具有促进生长发育、增强免疫力等生理功能。其天然、安全、多功能的优势,使其在婴幼儿配方奶粉、运动营养、医疗营养、乳制品及烘焙食品等领域得到广泛应用。随着消费者对健康营养和高蛋白食品需求的增长,全球乳蛋白市场持续扩大。根据Mordor Intelligence统计,2025年,全球乳蛋白市场产值约为13.8亿美元(约合人民币99.0亿元),预计到2030年将增至18亿美元(约合人民币129亿元),年均复合增长率约5.52%[89]。
4.1 乳铁蛋白
乳铁蛋白是一种非血红素类铁结合糖蛋白,包含703个氨基酸,分子量为80 kDa,广泛分布于乳汁、血清、唾液及其他外分泌体液中。乳铁蛋白约占母乳中总蛋白质的15%~20%,在初乳中的浓度尤为丰富,平均可达5~6 mg/mL[90]。其在调控新生儿先天免疫系统中发挥重要作用,是婴儿抵御外界病原体侵袭的首道免疫屏障[91]。乳铁蛋白的生物功能广泛,除参与机体铁代谢调节、维持铁稳态外,还具备抗菌、抗病毒、抗寄生虫、抗过敏、抗肿瘤及抗氧化等多种生理活性[92]。目前,乳铁蛋白主要通过牛乳提取获得,但天然含量极低,提取效率受限。每14 kg牛乳仅可提取约1 g乳铁蛋白,且受限于畜牧养殖用地紧张、资源消耗高等因素,我国乳铁蛋白高度依赖进口,对外依存度高达98%。与传统畜牧养殖方式相比,微生物合成可显著降低56%的CO2排放和87%的土地占用,是一种具有较强竞争力的经济、高效、绿色的乳蛋白合成方式[91](图6)。
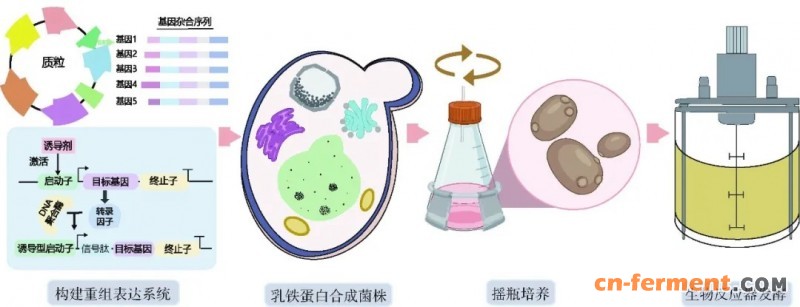
图6 乳铁蛋白微生物合成途径
近年来,得益于合成生物学与发酵工程的进步,乳铁蛋白的微生物合成逐步实现产业化突破。TurtleTree、All G和NewPro等企业构建了高效表达系统,大幅提升了产率并显著降低成本。研究者早期构建了乳铁蛋白原核表达载体pET-32a-LF,在E. coli中实现了乳铁蛋白的可溶性表达,经优化诱导条件后,乳铁蛋白表达量可达15.3 mg/L [93]。近年来,P. pastoris成为乳铁蛋白合成的重要宿主。2023年,Lv等[94]开发了一种将空间分离的CO2热催化与酵母发酵相结合的化学生物级联催化(CBCC)系统,实现了CO2向乳铁蛋白的高效转化。通过突破限速步骤,研究者构建了可高效合成人乳铁蛋白(hLF)的毕赤酵母菌株,在生物反应器中实现了5.4 g/L的产量,为已报道的最高产量。同时,NewPro公司也采用P. pastoris底盘进行乳铁蛋白表达,进一步提高了生产效率。此外,NewPro公司还探索了多种丝状真菌作为乳铁蛋白合成的底盘,丰富了微生物合成的宿主选择。这些研究不断推进着乳铁蛋白微生物合成技术的发展,提高了产量和生产效率,为其在食品、医药等领域的广泛应用提供了有力支持。
4.2 骨桥蛋白
骨桥蛋白(osteopontin, OPN),又称唾液酸糖蛋白,是一种高度磷酸化并具甘露糖糖基化修饰的酸性糖蛋白,广泛存在于多种哺乳动物组织及体液中,由多种细胞如成骨细胞、免疫细胞及乳腺上皮细胞等合成和分泌。研究表明,OPN通过调控MAPK13、CCNE1、NF-κB等信号通路,参与细胞增殖、分化及免疫应答,对维持组织稳态与调节炎症反应具有重要作用。作为一种重要的细胞外基质蛋白,OPN在骨重塑、肿瘤转移、免疫调控等多个生理过程中发挥关键功能[95]。在母乳中,OPN是促进新生儿发育的重要活性成分,特别在肠道发育、免疫系统成熟、神经系统完善及骨骼形成中发挥多重积极作用。母乳中OPN含量远高于牛乳,且其糖基化修饰和生物活性更为复杂,因此成为乳源功能蛋白研究的重点对象。然而,由于天然乳液中OPN含量极低,传统提取方式难以满足规模化生产需求。
随着合成生物学的发展,微生物表达系统成为实现OPN高效生产的理想路径。英国合成生物学公司Better Dairy率先构建了基于乳酸克鲁维酵母(Kluyveromyces lactis)的工程菌株,成功实现了OPN的异源表达,并推进至中试放大阶段,为其产业化应用奠定了基础。同时,加州大学研究团队于2017年利用莱茵衣藻(Chlamydomonas reinhardtii)成功表达磷酸化修饰但未糖基化修饰的牛源OPN,并在动物实验中证实其具备与天然蛋白相似的生物功能。2025年,Zhang等[96]使用PAOX1启动子和混合信号肽PROSCW10-α来驱动人源OPN的表达和分泌,在P. pastoris中成功实现人源OPN (hOPN)的高效表达,进一步通过共表达转录因子、翻译因子和分子伴侣等策略来提高hOPN的产量,最终人源OPN在摇瓶发酵和生物反应器中的产量分别达到了35.6 mg/L和128.5 mg/L,为目前已报道的最高胞外表达水平。
5 挑战与展望
合成生物学等生物制造技术的快速发展,为功能食品蛋白的绿色化、经济化与高效化生产提供了新机遇。然而,我国功能食品蛋白生物制造产业的发展仍面临诸多挑战。首要挑战在于现有食品酶的特异性、催化活性和稳定性尚待提升。例如,TGase在工业应用中仍存在热稳定性差和交联效率低等问题。近年来,人工智能技术和酶从头设计技术的快速发展为解决该问题提供了新工具。人工智能技术能够高效揭示酶“序列-结构-功能”间的内在关联,从而赋能酶蛋白的从头设计或改造,以获得满足特定需求的定制酶。
其次,部分真核来源的功能食品蛋白在微生物宿主中的表达与分泌效率依然偏低。以骨桥蛋白为例,目前报道的最高产量仅为128.5 mg/L,难以满足工业化生产要求。造成该现象的主要原因有3个:1)一些大分子真核蛋白在微生物体系中难以正确折叠,易在胞内形成无活性的包涵体;2)微生物通常缺乏异源蛋白的特异性转运机制,导致其难以高效分泌至胞外;3)微生物分泌的蛋白酶可降解异源蛋白。因此,未来的研究可聚焦于开发高效的分子伴侣以促进蛋白正确折叠;重塑微生物细胞的转运与分泌系统以提升蛋白分泌效率;以及敲除内源蛋白酶基因以防止目标蛋白降解。
此外,重组功能食品蛋白的结构与功能可能与天然蛋白存在一定差异。这主要是由于不同细胞具有各自独特的蛋白质翻译后修饰系统,尤其是糖基化修饰途径。例如,在毕赤酵母表达系统中,其固有的N-糖基化修饰机制可能会导致重组蛋白发生过度甘露糖基化,形成高甘露糖型N-聚糖结构。这种结构不仅可能降低蛋白的催化活性与体内半衰期,还会增加其免疫原性。现有研究主要通过敲除毕赤酵母中的N-糖基化途径来抑制重组蛋白的过度甘露糖基化。然而,此类遗传改造往往会降低细胞的比生长速率和最大生物量,进而削弱重组蛋白的整体合成效率。未来研究可采用适应性进化技术,定向筛选获得生长性能回复的突变菌株,从而提升重组蛋白的生产效率。
功能食品蛋白的生物制造方式与传统制造方式存在本质差异,亟须建立与之配套的质量评价体系,包括检测指标与控制标准。当前,国际上已批准多款合成生物学来源的功能食品蛋白上市,相比之下,我国在该领域的监管审批进度滞后,面临国内市场被抢占的风险。为此,国内科研界应加速对生产工艺中高风险环节的安全性评估;同时,产业界需联合学术机构,率先构建团体标准与技术指南,为国家层面制定科学、规范的监管标准奠定基础。
参考文献:略
引用格式:刘龙. 功能食品蛋白生物制造的发展现状与挑战[J]. 食品科学技术学报,2025,43(4):1-14.LIU Long. Development status and challenges of functional food protein biomanufacturing[J]. Journal of Food Science and Technology, 2025,43(4):1-14.基金项目: 国家自然科学基金资助项目(32270096)。Foundation: National Natural Science Foundation of China (32270096).