近日,山东大学祁庆生/侯进研究团队于期刊J. Agric. Food. Chem.发表研究综述metabolic Engineering for Squalene Production: Advances and Perspectives,总结了利用微生物细胞工厂生产角鲨烯的最新进展,并全面概述了各种已实施的代谢工程策略和潜在的调控策略,以增强角鲨烯的生产,为角鲨烯和相关萜类化合物的进一步代谢工程提供参考。
微生物生产角鲨烯的研究进展
角鲨烯,连同许多其他的三萜和类固醇,从由IPP和DMAPP聚合产生的共同前体FPP天然合成。在真核生物中,IPP和DMAPP的生成主要通过MVA途径,而在原核生物中,则通过MEP途径。DXS酶在MEP途径中催化甘油醛-3-磷酸和丙酮酸合成DXP,随后由DXR酶催化生成MEP,MEP经过一系列酶促反应生成IPP和DMAPP。而MVA途径则从乙酰辅酶A开始,形成HMG-CoA,然后转化为MVA,最终生成IPP,IPP再转化为DMAPP。
由法尼基焦磷酸合酶(ERG20)合成的法尼基焦磷酸(FPP),通过角鲨烯合酶(ERG9)转化为角鲨烯。角鲨烯合成每分子需6分子IPP和13分子NADPH,产生36分子NADH,消耗18分子乙酰辅酶A和9分子葡萄糖。因此,酵母中角鲨烯的理论产量为0.226g/g葡萄糖。
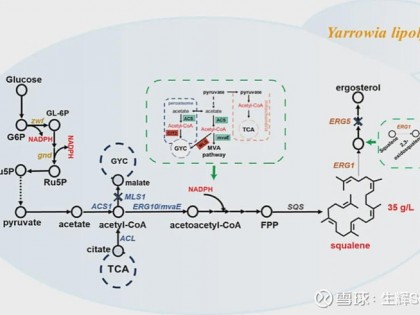
多种微生物已被改造用于生物合成角鲨烯。例如,通过过表达角鲨烯合酶和敲除竞争途径的关键酶,谷氨酸棒状杆菌可产生1.406g/L角鲨烯;集胞藻通过移除角鲨烯霍烯环化酶(SHC)可产生5.1mg/L角鲨烯;大肠杆菌通过代谢工程和细胞膜扩展被设计成可产生约612mg/L角鲨烯;裂殖壶菌通过优化培养条件和增强关键基因表达,角鲨烯合成增加了6倍,达到188.6mg/L;酵母中,过氧化物酶体工程被证明是提高角鲨烯产量的有效方法,如酿酒酵母中引入整个角鲨烯生物合成途径后,角鲨烯滴度达到11g/L;解脂耶氏酵母通过过氧化物酶体和细胞质工程实现了32.8g/L和51.2g/L的角鲨烯生产,这是目前微生物中最高的产量。
提高角鲨烯产量的代谢途径改造
通常,用于合成角鲨烯的原核生物最多产生约1g/L角鲨烯,而真核生物可产生约50g/L角鲨烯。目前,利用酵母细胞工厂生产角鲨烯已取得显著进展,因此,本综述主要总结酵母细胞工厂中使用的工程策略。
增强乙酰辅酶A供应
乙酰辅酶A是合成IPP的前体,在TCA循环和类固醇合成中起重要作用。在酵母中,通过PDC、ALD和ACS酶将丙酮酸转化为乙酰辅酶A。过表达这些限速酶,尤其是ACS2,能显著提高角鲨烯产量。在解脂耶氏酵母中,ACS和ACL酶负责生成乙酰辅酶A。通过引入异源ACS基因和过表达ACL2,角鲨烯产量增加了20%。此外,通过引入PK-PTA途径和乙醇转化为乙酰辅酶A的途径,也成功提高了乙酰辅酶A的供应。在酵母中,胞质MVA途径无法直接利用过氧化物酶体中的乙酰辅酶A。通过将CAT2与PEX15融合,增加了乙酰辅酶A的浓度,提高了角鲨烯产量。
辅因子工程
每合成一个角鲨烯分子需要13个NADPH分子。为了提高 NADPH 的可用性,通过增加G6P脱氢酶和6PD的表达,角鲨烯产量提高了104%。共表达NADH激酶基因POS5和tHMG1,角鲨烯合成提高了27.5倍。此外,表达NADH依赖的HMGR减少了对NADPH的依赖,而ANT1的过表达增加了过氧化物酶体内的ATP,使角鲨烯生物合成提高了27%。这些策略通过优化辅因子供应和能量利用,有效增强了角鲨烯的生产。
加强MVA途径
HMGR是MVA途径中的关键限速酶,通过N端截短的HMGR(tHMGR)可以显著增加MVA途径的流量。在酿酒酵母中过表达带有C末端催化域的tHMG1基因,使角鲨烯生物合成提高了40倍。通过高表达ERG13、HMG1和IDI等基因,以及引入整个角鲨烯生物合成途径到细胞器中,角鲨烯产量显著提升,解脂耶氏酵母中达到51.2g/L,酿酒酵母中达到21.1g/L。这些策略展示了通过增强MVA途径来提高角鲨烯产量的潜力。
抑制竞争途径
由于角鲨烯及其衍生物对细胞生长至关重要,完全阻断类固醇合成途径是不可行的。通过下调ERG1基因表达,可以减少角鲨烯代谢,增加其积累。例如,将ERG1启动子替换为PHXT1可提高角鲨烯产量11.3%至12.5%。进一步替换为较弱的PTHI1启动子,角鲨烯产量增加了20倍。ERG1突变体与弱ACL启动子结合也提高了角鲨烯生物合成。此外,化学抑制剂特比萘芬通过抑制ERG1,显著促进了角鲨烯积累,产量增加了最多35倍。这些方法展示了通过调控ERG1来增强角鲨烯生产的潜力。
细胞器工程
细胞器工程是提高萜类化合物合成的有效方法,包括隔离合成途径、规避复杂调控机制,并提供丰富的乙酰辅酶A和辅因子。通过将整个角鲨烯生物合成途径引入过氧化物酶体,可显著提高角鲨烯产量。此外,线粒体作为乙酰辅酶A和ATP的储存库,其工程化也用于提高萜类化合物产量;内质网的扩张通过INO2过表达增加了角鲨烯和原人参二醇的产量;细胞壁的稳定性对萜类化合物的合成至关重要,移除ECM33可以提高膜的稳定性,增加角鲨烯合成。
引入合成代谢途径生产IPP和DMAPP
天然MVA途径效率低下且复杂,为解决这些问题,研究创建了异戊烯醇利用途径(IUP)。IUP途径步骤简单,只需两步反应即可将异戊烯醇或戊烯醇转化为IPP和DMAPP。尽管IUP途径具有简单和减少催化步骤的优势,但它也需要外部物质,这可能导致细胞毒性和增加生产成本。
提高萜类化合物产量的代谢途径调控
除了探索促进角鲨烯合成的传统策略外,研究还讨论了增强其他萜类化合物合成的各种方法,特别是脂溶性萜类化合物。其中一些策略,如DGA1和ACC1的过表达,已经应用于角鲨烯合成,而其他策略,如动态调控,尚未被利用。总体而言,这些策略为未来改善角鲨烯的生物合成提供了宝贵的见解。
脂质合成调节
角鲨烯作为一种疏水分子,主要分布在脂质体和细胞膜中。可通过增强TAG合成来提升酵母中萜类化合物的积累,DGA1基因编码的酶在TAG合成中起关键作用。DGA1和HMG1共同表达可显著增加角鲨烯产量。过表达PAH1、DGA1和ACC1可增强TAG合成,提高番茄红素产量。解脂耶氏酵母能利用油脂作为碳源,通过过表达DGA1和CAT2基因,可从废油中合成角鲨烯。减少β-氧化可增加脂质含量,促进萜类化合物储存。适当脂质水平对萜类化合物生产至关重要,过度合成可能导致碳源和能量耗尽。因此,增强脂质合成以储存更多角鲨烯是代谢工程中的重要策略。
转录因子调控
角鲨烯是麦角固醇的前体,对真菌细胞膜的柔韧性、穿透性和膜相关蛋白的功能至关重要。在酿酒酵母中,麦角固醇合成基因的调控涉及UPC2和ECM22等类固醇调节元件结合蛋白,以及MOT3和ROX1等抑制因子。UPC2转录因子在类固醇水平低时通过类固醇调节元件激活类固醇生物合成基因的表达。UPC2的突变体UPC2-1能够使细胞在有氧条件下吸收类固醇,增强激活域的效果,从而显著增加 amorpha-4,11-二烯和角鲨烯的产量。
提高不溶性萜类化合物分泌
由于角鲨烯的疏水性和大分子尺寸,它难以穿过细胞膜,导致在细胞内积累和随后的代谢负担。为此,有研究设计了一种合成途径,使用带有信号肽的角鲨烯结合蛋白因子(SPF)来装载和运输角鲨烯,显著增强了角鲨烯的分泌,产量达到226 mg/L;还有利用ATP结合盒转运蛋白PDR5和氧化固醇结合同源蛋白3(OSH3)来增强角鲨烯和β-胡萝卜素的外排,这些进展为减轻细胞内代谢物积累引起的代谢压力提供了新思路。
酵母的代谢工程,包括增加前体供应、工程辅因子、加强MVA途径、抑制竞争途径等,已广泛应用于萜类化合物的生产。亚细胞工程,如线粒体或过氧化物酶体工程,结合细胞质工程,已实现克级角鲨烯生产,显示出提高萜类化合物产量的巨大潜力。除了工程化与萜类化合物合成直接相关的基因外,还可以识别新的基因靶点以提高萜类化合物产量。动态调控也被证明是提高萜类化合物产量的有效方法。未来应用这些策略不仅将进一步增强角鲨烯生产,还将为萜类化合物的代谢工程提供宝贵的洞见。
文献来源:
https://doi.org/10.1021/acs.jaf