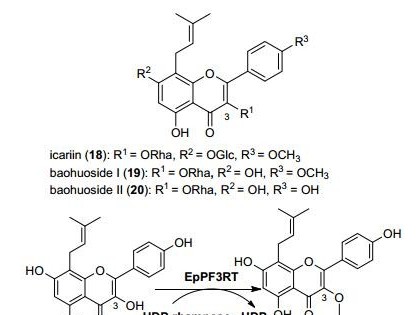
随着人们膳食结构的改善、环保意识的增强,加之饲料资源匮乏的严峻现实,大麦、小麦、次粉、糠麸等替代玉米,菜粕、棉粕、花生粕等替代鱼粉和豆粕成为普遍,但其中的抗营养因子如木聚糖等限制了其在饲料中的有效应用,因此饲料酶制剂特别是木聚糖酶成为近年研究的热点。
1 木聚糖结构与抗营养作用
小麦、玉米、豆粕、杂粕等是目前饲料行业中常用到的饲料原料,其营养价值的差别主要是:①蛋白及碳水化合物在数量和质量上的差异;②内源性酶活上的差异。谷物类饲料如玉米、小麦、大麦等,一般由果皮、种皮、糊粉层、次糊粉层和含淀粉胚乳组成。其中淀粉是易消化的营养性多糖,纤维素、半纤维素、果胶和抗性淀粉(阿拉伯木聚糖、β-葡聚糖、甘露聚糖、葡糖甘露聚糖等)构成植物籽实细胞壁的结构多糖和基质多糖,称为非淀粉多糖(None Starch polysaecha-rides NSP),因糖苷键与淀粉的不同,不能被动物内源酶分解。
细胞壁维持着植物的结构完整性,木质素为苯丙烷衍生物的聚合物(亦称苯丙基的多聚物),通常与细胞壁中的多糖形成动物内源酶难降解的复合物,限制动物对植物细胞壁物质的利用。纤维素作为细胞壁坚硬的骨架结构浸埋在半纤维素、果胶等基质多糖和糖蛋白构成的无定型胶状基质中,为细胞壁的结构多糖;果胶与纤维素、半纤维素结合形成不溶性的原果胶(见图1)。
半纤维素和纤维素是两种完全不同的物质(如表2)。半纤维素的成分是杂聚多糖,通常按其组成主链的占多数的糖来命名,被称之为木聚糖、甘露聚糖、葡甘露聚糖和半乳葡甘露聚糖等。主链的连结键为β-1,4键,许多支链又称侧链上的糖类有阿拉伯糖、半乳糖、甲基葡糖醛酸。
木聚糖是半纤维素的一种,属植物细胞壁中的结构性非淀粉多糖,在植物细胞壁中的含量仅次于纤维素,约占细胞干重的35%。它是由D-木糖单位主干[以β(1-4)键相连)和L-阿拉伯糖分枝[以α(1-2)键和α(1-3)键相连]所组成的吡喃型木糖聚合物,又称阿拉伯木聚糖或戊聚糖。阿拉伯糖的取代降低了主链化学键的作用力,从而使其具有水溶性和粘稠性。
试验证明,木聚糖能结合大量水分(Patel等,1980;Philp,1995),小麦中阿拉伯木聚糖可吸收约10倍于自身重的水,增加消化道内容物和粘稠度,是谷物中的重要抗营养因子。其危害:
①形成营养屏障,降低溶质的扩散速度(Petterson等,1989)阻碍养分和内源酶的作用,降低蛋白质、氨基酸、脂肪等营养物质的消化吸收和利用。进一步研究表明,木聚糖能直接结合消化道中的蛋白酶、脂肪酶等消化酶,使其活性降低。由于代谢补偿作用,增加了蛋白质、脂类和电解质等内源物质的分泌,降低它们在体内的储备(Irish,1993;Campbell,1983,1986等)。
②排空速度减慢,促进后肠道微生物发酵,养分利用率降低。
③影响消化道生理形态、内分泌及胃肠道激素水平,降低动物生产性能(Isaksson等,1982;Ikegami等,1990;于旭华,2001)。
④降低家禽和猪的表观代谢能。木聚糖含量直接影响到饲料的表观代谢能值,Choct等(1990)检测了NSP与表观代谢能的关系发现,随着NSP含量的增加,饲料表观代谢能呈线性降低。Annison(1993)试验不同的小麦品种因其阿拉伯木聚糖含量的不同,对鸡表观代谢能的影响范围为10.4-15.9MJ/kg干物质。
2 木聚糖酶的结构和酶学性质
2.1 木聚糖酶的来源
木聚糖酶是一类将木聚糖降解成低聚糖和木糖的一种复合酶,许多植物(如大麦、稻谷等)、细菌及少数真菌(如木霉、曲霉等)都可产生混合纤维弧菌(Cdlvibrio mixtus)和荧光假单胞菌(Pseudomonas fluorescens),也可产生内切β-1,4-木聚糖酶。研究发现:能产生木聚糖酶的微生物至少有20种,包括厌氧菌(Anaerobes)、需氧菌(Acrobes)、嗜温微生物(Thermophiles)、嗜热微生物(Thermophiles)和极端微生物(Extremophiles)等。各种来源的木聚糖酶已经通过产业化发酵生产,并在饲料中得到广泛应用。
2.2 木聚糖酶的结构
木聚糖酶是具有四级结构的大分子活性蛋白,在结构上有催化结构域、底物结合结构域、热稳定结构域、连接区域、Cellulosome-Docking结构域、重复序列和其它未知功能的非催化结构域。不同木聚糖酶含有其中的一个或一个以上的区域。来源不同的木聚糖酶氨基酸组成相差不大,主要为丙氨酸、谷氨酸、甘氨酸、丝氨酸和苏氨酸等。但谷氨酸、天门冬氨酸、酪氨酸、色氨酸、甘氨酸和丝氨酸对其催化特性及耐碱、耐热性等非常关键。
细菌产生的木聚糖酶蛋白质亚基比较单一,真菌产生的木聚糖酶蛋白质亚基则较复杂,分子量大小变化也较大。以氨基酸序列相似性和疏水簇分析法为基础,Henrissat和Bairoch(1993,1996)将糖基糖苷键水解酶中的内切木聚糖酶分为10族和11族,它们分别组成高分子量(>30kD)和低分子量(<30kD)的木聚糖酶。10族内切木聚糖酶主要包括酸性和大分子的内切木聚糖酶,而碱性的和小分子的内切木聚糖酶则属于11族。通常细菌可产生两种木聚糖酶:低分子量的碱性木聚糖酶和高分子量的酸性木聚糖酶。真菌只产生低分子量的碱性木聚糖酶。
2.3 木聚糖酶的酶学性质
木聚糖酶分子量范围为8-145kD,等电点在3-10之间,pH范围为3-10,多数酶的最适pH范围为4-7,在pH2.5-12范围内酶活性能保持80%。最适温度为40-60℃,60℃时酶活保持在80%,以后随温度升高而下降。Cu2+、Zn2+、Pe2+、Pe3+对木聚糖酶活性有抑制作用,Co2+、Mn2+对其有促进作用。一般细菌所产木聚糖酶比真菌所产木聚糖酶热稳定性好,真菌中也有个别耐高温的木聚糖酶。
2.4 木聚糖酶的作用方式
木聚糖酶作用方式是典型的酸碱亲核水解反应。作用于木聚糖主链的酶有三种:内切β-1,4-木聚糖酶(EC 3.2.1.8)、外切β-1,4-木聚糖酶(EC 3.2.1.92)和β-木糖苷酶(1,4-β-D-木聚糖木糖水解酶EC3.2.1.37)。内切酶从主链内部作用于木糖苷键,将木聚糖分解成低聚糖,使黏度降解,是关键酶,外切酶只能在主链末段发挥作用,降低粘性作用较弱,β-木糖苷酶则作用于低聚木糖的末端,释放木糖,在消除抗营养作用方面意义不大。彻底水解木聚糖取代基支链还需要乙酰酯酶、α-L-阿拉伯糖酶、α-一葡糖醛酸酶和一木糖苷通透酶等特异性酶。
内切酶又可分为特异性和非特异性木酶两种(Coughlan,1992),特异性木糖内切酶仅对木糖的β-1,4键有活性,非特异性木糖内切酶可水解以β-1,4键连接的木糖、混合木糖的β-1,4键及其它以β-1,4键连接的多糖。大多数内切木糖酶能特异性地作用于木聚糖的非取代木糖苷键,并释放取代的和非取代的木寡糖,而其它内切木糖酶特异性作用于主链上的接近取代基团的木糖苷键。
一种微生物常常产生不止一种类型的木聚糖酶,本质上都有内切酶和外切酶,只是活性高低不同而已。由真菌发酵所产木聚糖酶大多以外切酶为主,以细菌发酵产生的木聚糖酶以内切酶活性为主。
3 木聚糖酶在饲料中的应用
饲料中一般含有较多的非淀粉多糖(Nonst-arch Polysaccharides,NSP),主要包括阿拉伯木聚糖、β-葡聚糖、α-半乳糖苷、果胶、纤维素等。其中前两者占NSP的30%。NSP分为可溶性NSP(SNSP)和不溶性NSP(INSP),影响动物对养分进行消化的主要是SNSP,β-葡聚糖和木聚糖是两种较为重要的SNSP。
3.1 木聚糖酶在猪饲料中的应用
事实表明,饲料中添加β-葡聚糖酶和木聚糖酶在猪和家禽中的饲喂效果是不一致的。原因可能是:
3.1.1 消化环境不同
低pH值、胃蛋白酶和胰酶均可使外源酶活性降低(Inborr等,1993;Almirall等,1995b),畜禽消化道中pH不同对酶活的影响最大(史凯来等,2000)。饲料在猪相对较低的pH条件下要停留4-12h,而在鸡低pH的前胃中仅仅停留20-40min。Thacker等(1996)和Bass等(1996)分别用5种酶做了阿拉伯木聚糖酶和β-葡聚糖酶在猪的pH值模拟试验,表明木聚糖酶长时间处于较低的pH值条件下能够破坏其中的一部分酶活。进一步做的猪小肠食糜中酶活测试发现,酶在经历了低酸过程后小肠中的活性仍然保持很高(在pH值2.5的条件下分别保持15min和120min后pH值升至5.5,木聚糖酶的酶活分别降为原酶活的12%-79%和12%-48%),说明尽管体内胃蛋白酶可能加速酶的失活,而饲料中的其它成分对酶可能具一定的保护作用。
3.1.2 消化方式不同
猪的小肠可以降解饲料中大量的β-葡聚糖和木聚糖。其对木聚糖的降解程度较低,但整个消化系统对木聚糖的降解率也达65%(Weltzien等,1986),作用结果是小肠微生物作用还是饲料内源酶尚无定论。
3.1.3 小肠内容物粘度不同
肉鸡小肠内容物粘度大约是猪的100倍。猪小肠内容物的水分含量(10%的干物质)高于肉鸡小肠内容物的水分含量(20%的干物质),因而降低了可溶性碳水化合物的粘性。Bedford(1991)试验酶对猪小肠内容物的粘性没有显著影响。
饲用酶应用于猪日粮中原因有三:(1)减少仔猪断奶应激;(2)提高饲料的利用率;(3)降低环境污染。添加木聚糖酶能提高猪对高纤维饲料的利用率,减少仔猪腹泻率(Bedford,1995;Imborr和Oyle,1998)。猪日增重改进的范围为4.5%-5%,饲料转化率改进的范围为3%-15%,其中报导饲料转化率改进的情况比日增重多。添加木聚糖酶一般对玉米-豆粕型日粮效果不明显,较多的试验证明在大麦、小麦或黑麦为主日粮中有明显的正效应。但Thacker等(1991、1992)报道在小麦、大麦、燕麦日粮中加木聚糖酶也无效果。
白亮亮(1999),程伟(1998),冯定远(1997)报道,在20-60kg生长育肥猪的次粉或小麦日粮中添加5%木聚糖酶制剂可提高日增重和饲料转化率,降低腹泻频率,健康状况明显改善。冯定远等(1998)在断奶仔猪日粮中使用β-葡聚糖酶和木聚糖酶,使实验组生长仔猪日增重提高6.0%。许梓荣(1999)、王修启等(2000)也得到了同样的结果。张克刚等(1997)在含有高量小麦麸和次粉的育肥猪日粮中使用酶制剂,实验组日增重分别比对照组提高4.25%和14.02%,高量次粉组比高量麦麸组效果更好。Prokopp(1999)报道,木聚糖酶和蛋白酶制剂应用于仔猪,能提高仔猪日增重,降低仔猪死亡率,且能使蛋白质消化率提高3%,氨基酸浓度提高3%-5%(P<0.01),纤维消化率提高17%(P<0.01),氮利用率提高10%(P<0.01),蛋白质沉积率提高9%(P<0.01)。Schohen等(1997)报道,在断奶仔猪饲以小麦与麦麸混合日粮中添加木聚糖酶和β-葡聚糖酶,断奶后前3周酶的添加对死亡率无影响,3周后腹泻率有所降低。
王冬艳等(2000)饲养试验表明:对断奶仔猪,复合酶制剂(蛋白酶、木聚糖酶等)对生长性能和腹泻的影响比单一纤维素酶制剂要好。奚刚(1999)研究证实,饲料添加外源性酶能刺激某些内源消化酶的分泌,与Inborr认为的外源性酶有利于动物的内源性酶,不存在“反馈性抑制”的结论一致。高峰(2002)、许梓荣(1999)研究发现:木聚糖酶使仔猪血糖浓度升高,机体免疫增强。陈华清(2005)试验木聚糖酶使仔猪的甲状腺素T4含量显著升高,T3也有提高,促进胃排空、内源酶分泌和胃肠道上皮增生的胃泌素分泌增强,一定程度上提高了猪的采食量,加强了体液调节和营养物质的代谢。
3.2 木聚糖酶在鸡饲料中的应用
木聚糖酶可以提高不同小麦品种鸡的表观代谢能、养分消化率和氨基酸的消化率,显著降低空肠和回肠的食糜粘度,有效拆除营养吸收屏障,消除阿拉伯木聚糖的抗营养作用,加强对营养物质的吸收利用。Ollofs-k等(2000)报道,小麦日粮添加木聚糖酶可以提高蛋鸡生产性能,并认为其原因是木聚糖酶提高了小麦日粮蛋鸡的表观代谢能和降低肠道食糜粘度所致。林东康(1999)用小麦替代日粮中30%的玉米饲喂蛋鸡(粉料),结果表明,酶制剂提高饲料转化率2.9%-3.3%,产蛋率提高1.3%-2.1%,但统计检验差异不显著。王修启试验木聚糖酶使蛋鸡产蛋率提高6.17%。总蛋重提高11.73%,平均蛋重提高5.24%,料蛋比降低了11.30%,差异均达到极显著水平(P<0.01)。
另外,酶制剂除了直接作用机制(提高营养物质的消化率)以外,还通过改变肠道的菌群数量和细菌种类,利用肠道微生物菌群的互作来改善动物生产性能。王金全等(2005)在AA肉仔鸡分别饲喂玉米-豆粕型,小麦-豆粕型和小麦-豆粕型添加木聚糖酶3种日粮的试验表明,除需氧菌之外,小麦NSP显著增加了回肠厌氧菌总数(P<0.05),其他菌类(乳酸菌、大肠杆菌、粪链球菌)只表现出增加趋势,加酶后需氧菌、乳酸菌、粪链球菌的数量增加,厌氧菌、大肠杆菌的数量降低,有利于物质的再消化吸收。
酶制剂可以提高肉鸡的免疫力(韩正康,1996)。王金全(2005)、崔朝霞等(2003)试验证明,木聚糖酶降低肠道食糜黏度(P<0.05),使肉仔鸡的脾脏、胸腺和法氏囊的相对重量均提高(P>0.05),死淘率下降60%(P<0.05)。同时能降低胰脏、肝脏、小肠、肌胃和腺胃消化器官的相对重量(P>0.05)。血液代谢激素结果表明,添加木聚糖酶对T3、GH、INS激素水平表现出增高趋势(P>0.05),而T4、TSH和GLU水平加酶后有所降低(P>0.05)。
Steenfeldt et al.(1998)、Choct(1995)、高峰等(2000)、王修启(2005)研究,小麦基础日粮添加木聚糖酶使肉鸡平均日增重提高、料重比降低。日粮添加木聚糖酶可显著提高葡萄糖的吸收,木聚糖酶主要作用在小肠上段,并且伴随小肠上段SGLTI表达的上调。
3.3 木聚糖酶在鸭鹅饲料中的应用
酶制剂可改善鸭的生产性能,增加动物肝重、降低小肠重量(张彬等,2000),提高种番鸭的产蛋量和受精率(孙淑霞等,2001)。应用方式有两种:一是在正常营养水平的日粮中添加酶制剂;二是在营养水平降低的日粮中添加酶制剂。但何健试验低营养水平(降低12%)日粮添加酶制剂对3-4周龄阶段肉鸭效果显著,对1-2周阶段肉鸭有不良影响,应慎重使用。张堂田等(1999)在玉米-豆粕型日粮中添加木聚糖酶、β-葡聚糖酶、纤维素酶可显著提高樱桃谷肉鸭的生产性能,木聚糖酶是影响肉鸭生产性能的最主要的因素。高树冬等(2001)以不同比例的小麦添加不同比例的酶制剂,替代等量的玉米,结果发现,可提高饲料转化率2.26-4.53%,提高成活率10.73-12.02%,降低饲料成本4.98-10.96%。朱元招等(1999)在玉米-豆粕-麦麸型日粮中添加两种复合酶制剂研究其对樱桃谷肉鸭生产性能的影响,发现木聚糖酶、蛋白酶、果胶酶组合饲喂效果比淀粉酶、木聚糖酶、蛋白酶组合好。徐欢根等(2001)的饲养结果表明:用添加酶制剂的小麦-豆粕型日粮饲喂绍兴麻鸭,其增重、料肉比、产肉与饲料成本明显改善。很多试验表明,添加外源酶制剂可适当增加杂粕在饲粮中的比例,提高鸭的体增重、饲料转化率、蛋白质消化率,降低日粮成本(李昂等2004,2005)。高宁国(1997)等报道,在以大麦为主的肉鸭饲粮中,添加β-葡聚糖酶为主的酶制剂,21日龄时,使食糜上清液的淀粉酶活力提高了75%。杨桂芹等(2005)试验表明:二指肠食糜中蛋白酶活力增大,提示木聚糖酶在解除木聚糖抗营养作用的同时饲料中的蛋白质与消化酶相互作用加强,提高了饲料蛋白质的消化率,加木聚糖酶可使肉鹅淀粉酶和蛋白酶活性升高,当添加量为0.1%时,肉鹅内源淀粉酶活性最高,比小麦组高出20.57%,比玉米组高出12.05%;当添加量为0.2%时,肉鹅蛋白酶活性最高,比小麦组高出34.48%,比玉米组高出29.40%。
4 木聚糖酶研究进展
木聚糖酶的研究主要集中在酶学性质和实际应用中的障碍。现在,为提高饲料的利用率、适口性和市场竞争力,饲料进行二次制粒或膨化成为普遍。酶的稳定性受到了挑战。首先从耐热性的微生物体内分离生产耐热性的酶类,Pettersson和Rasmussen(1997)研究证实,由高温霉(The Flnomyces)、腐质霉(Humicola)和木霉(Trichoderma)分离到的木聚糖酶在热稳定性上存在差异,由木霉(Trichoderma)分离到的木聚糖酶在75℃二次调质温度下明显失活。而由高温霉与腐质霉分离到的木聚糖酶在85℃二次调质温度下可保留80%以上的活性。其中由高温霉分离到的木聚糖酶,即使在95℃二次调质温度下仍可保留70%以上的活性。其次进行酶产品加工改造如制粒、包丸。第三是在制粒冷却后增加后喷涂酶环节。但这需要额外增加某些加工设备及加工工序(Perez-Vendrell等,1999b)。
利用生物反应器生产木聚糖酶,运用基因工程技术在分子水平上对木聚糖酶基因进行分子改造,可望解决一些木聚糖酶活性(如抗逆性、pH、热稳定性等)低的应用缺陷。自1983年以来,木聚糖酶基因的分子克隆和表达已取得很大进展,通过构建基因载体及特殊受体菌株重组手段成功地使一些木聚糖酶基因得到高效表达。目前,研究外源木聚糖酶基因的表达和调控机制的受体菌株主要为Ecoli。木聚糖酶基因分子克隆工作已把宿主菌从大肠杆菌属等原核生物扩展至酵母、木霉、曲霉等真核生物。
另外的途径是生产转基因饲料植物。科学已证实,外源β-葡聚糖酶和木聚糖酶在转基因植物中可以表达,且表达产物具有生物学活性,酶的表达大多情况下似乎不影响植物的生长发育,说明转基因植物具有可行性。非淀粉多糖酶转基因植物可作为生物反应器生产酶制剂,而更有效的方法是在玉米、大豆、大麦种子等饲料原料中表达非淀粉多糖酶,直接用作饲料,这样完全省去酶的生产和添加过程,取代目前通过微生物发酵生产酶制剂的方法。
5 木聚糖酶使用中的技术问题
近两年我国玉米、豆粕等原料价格的不断波动,使得小麦和各种杂粕的使用量增加,小麦酶和杂粕酶的使用已日益广泛,包括湖北、河南、河北、山东甚至东北各省份复合酶使用已非常普遍。另外,全球经济一体化,使国外低价格小麦、大麦产品不断涌入中国市场,加之国内陈化粮小麦的再利用现实,都将改变传统的饲料配方模式,进一步刺激木聚糖酶的市场需求。因此,针对市场需求酶制剂发展方向应集中在两方面:一是单项酶制剂的研究与开发。木聚糖酶要研究内切、外切酶有合适比例的“结合”型才能消除木聚糖的抗营养作用,适应于国内的传统日粮类型。二是根据不同日粮类型、不同动物种类以及不同抗营养因子特点开发专用“日粮酶”、“动物酶”等复合酶。如“玉米-豆粕”型专用酶、“小麦-豆粕”型专用酶、“玉米-小麦-豆粕”型专用酶、“玉米-杂粕”型专用酶。不同酶制剂配合使用,效果优于单一酶制剂(Piao等1998;李化平译,1991)。后者提供给饲料企业的方便、快捷、彻底服务和针对性越来越显示出其强劲优势。
发挥木聚糖酶的效能,首先要研究专一木聚糖酶在不同日粮中的添加配比与提供消化养分的当量关系。冯定远等(2000)报道,在猪日粮中加入木聚糖酶能使干物质消化率提高11.3%。黄金秀等(2006)试验回归分析表明:养分消化率与饲粮木聚糖含量间呈较强的线性回归关系,木聚糖含量每增加1个百分点,粗蛋白、有机物、Ca、P、ADF和能量的消化率分别下降0.94、0.73、4.07、1.94、3.34、0.76个百分点,体重为15公斤左右的仔猪对木聚糖具有较强的耐受力,随着木聚糖含量增加,饲粮养分的消化率呈不同幅度下降,当玉米-豆粕型日粮木聚糖含量低于7.25%时添加木酶对养分消化率无改进作用,达到或高于7.25%时才有一定改进作用。Choct等(1990)在比较包括小麦、黑麦、小黑麦、大麦、高粱、大米和玉米7种日粮的AME时发现,各种饲料原料的AME值与其中阿拉伯木聚糖含量之间存在着强的负相关关系,相关系数为0.95。尽管酶制剂在玉米-豆粕型日粮中的应用效果不如在麦类日粮中明显,甚至营养水平较高的玉米-豆粕型日粮中添加酶制剂没有明显效果(Charlton,1996),造成配方成本提高,使用酶制剂没有空间等。但添加酶制剂的潜力已在实践中得到了引证。Pack和Bedford(1997)、Pack等(1998)的研究表明:含有淀粉酶、木聚糖酶和蛋白酶的复合酶制剂对玉米-豆粕型日粮营养价值有一定作用,其中玉米的可利用能提高了2%-5%。程时军等(2006)比较了复合酶及木聚糖酶在肉鸡玉米-豆粕型日粮饲养中的差别试验,证明酶制剂可降低全期死亡率,降能加酶可增加收益2-3%。加酶100ppm的潜在营养价值相当于50KcaL/kg禽代谢能,饲养前期(0-3周)复合酶效果好于单一木聚糖酶,饲养后期(4-6周)木聚糖酶的改善效果好于复合酶。Stanley等(1996)报道,使用含有蛋白酶、阿拉伯木聚糖酶、纤维素酶、β-半乳糖苷酶和淀粉酶组成的复合酶均可明显提高肉鸡棉粕日粮的饲料转化效率,棉粕量分别为7.5%、15%和30%。Pack等(1996)报道,在肉鸡高粱(65%)、豆粕或高梁(62%)、豆粕、菜籽粕日粮中添加木聚糖酶等进行试验(菜籽粕前期5%、后期10%),结果表明,日粮中添加酶制剂提高了高梁日粮肉鸡全期的增重和饲料转化率,高粱豆粕日粮组加酶后达到显著水平(P<0.05)。表明酶制剂不仅可应用于玉米豆粕型日粮中,而且对高粱菜籽粕日粮同样有效(菜籽粕用量可达10%),未影响动物的生产性能。冯定远(2005)在综合有关试验报道的基础上,建立了常见植物能量和蛋白质饲料原料使用相应酶制剂的ENW(Effective Nutrients Improvement Value,ENIV)值,即“有效营养改进值”,量化指导配方计算,避免了酶制剂实践中的盲目性。
酶制剂包括木聚糖酶在畜禽饲料中科学使用需要做的工作还很多,如酶活标准的统一、酶活简捷快速的检测方法普及、各地饲料原料中抗营养因子的含量范围、单酶配伍的拮抗与促进关系、针对不同畜禽品种、不同生长阶段对应日粮类型添加酶制剂的营养数据模型体系等。